 カナダ発!
"たんぽっぽ"
自閉症療育支援の~と
Autism Resources
カナダ発!
"たんぽっぽ"
自閉症療育支援の~と
Autism Resources
東京大学は2月6日、脳の発達障害に関連する分子である「DCLK」が突起の先端に局在して突起形成を促進すること、それと同時にシナプス内にも入り込み、突起先端でシナプスの形成が過剰に起こることを抑制することを見出したと発表。
また、DCLKは「N末端」に「微小管結合ドメイン」、「C末端」にタンパク質のリン酸化を行う「キナーゼドメイン」を持つキメラ分子(由来が異なる複数の部分から構成されている分子)だが、N末端ドメインが突起を伸ばし、C末端ドメインがシナプス形成を阻害することで相反する作用が生じることも解明したことも発表した。
成果は、東大大学院 医学系研究科 神経細胞生物学分野の岡部繁男教授らの研究グループによるもの。研究の詳細な内容は、2月5日付けで英国科学雑誌「Nature Communications」オンライン版に掲載された。
脳の発達は、遺伝的要因と環境要因の相互作用によって影響を受けてしまう。特に脳の発達障害を引き起こす遺伝子にはさまざまなものがあり、それぞれの遺伝子の持つ役割を明らかにすることで、脳の正常機能と
病態を理解する道筋が拓けると考えられている。
例えば「滑脳症」と呼ばれる発達障害では、原因遺伝子である「Lis1」や「DCX」に異常があることで幼若な神経細胞が正しい場所に移動することができなくなり、その結果、脳のしわがなくなる「滑脳」が生じるという。
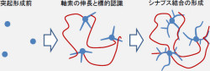
これまで、脳の発達障害向け研究としては、幼弱脳での細胞分裂や細胞移動を対象とした解析が主に行われてきたが、滑脳症の患者に見られる知的発達障害や脳の過剰興奮などの症状が、神経細胞の移動の障害に関連しているのか、神経回路の機能変化などの別の原因で起こっているのかについては不明のままであったほか、神経回路の形成のメカニズムの解明に向け、これまでさまざまな研究が行われてきたが、神経細胞が「樹状突起」と「軸索」の2種類の突起を作り、次に長く伸びた突起の間で接着が起こり、「シナプス」を形成する過程においては、突起の伸長とシナプスの形成は共に回路の形成を促進するが、実際には突起が十分伸びた後でシナプス形成を開始する必要があり、過剰なシナプスの形成が早期に起きてしまうことは正常な回路形成にとっては有害だと考えられるなど、未だ同定されていないさまざまな制御機構が存在することが考えられている。
また、滑脳症の原因遺伝子であるDCXと同じ遺伝子ファミリーに属するDCLKは、モデルマウスの解析からDCXと同様に幼若脳での神経細胞の移動に関与する分子であると考えられてきたが、その神経回路形成における役割は不明のままであった。
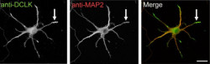
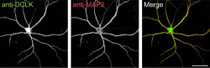
研究グループは今回の研究から、DCLKがシナプスを介して情報を受け取る構造である樹状突起の先端に局在すること、ならびに局在したDCLKが「細胞骨格」である微小管を束化することで突起形成を促進することを見出すことに成功したほか、DCLK分子がシナプス内にも存在し、シナプスの形成が過剰に起こることを
抑制したり、情報を受け取るシナプス後部に存在する「グルタミン酸受容体」の量を低下させることでシナプスを介した情報のやりとりを抑制することも見出した。
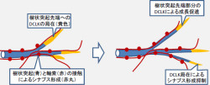
また、DCLKはN末端に微小管結合ドメイン、C末端にタンパク質のリン酸化を行うキナーゼドメインを持つキメラ分子であり、N末端のみ、あるいはC末端のみの変異型分子を神経細胞に発現させることで、N末端ドメインが樹状突起伸長を促進し、C末端ドメインがシナプス形成を阻害するといった回路形成に対して相反する2つの作用を担っていることも判明した。
さらに、DCLK分子の脳内での発現は出生直後に高く、脳の成熟に従って発現量は低下していくことが
確認されたことから、樹状突起の成長がほぼ終結
し、シナプス形成を抑制する必要がなくなるとDCLK
も役割を終えて量が減っていくという仕組みが考え
られると研究グループではコメントしている。
なお、今回の成果について研究グループは、脳における神経細胞同士のつながり方に対して、新しいモデルを提供するものとなるとしており、これにより遺伝性の脳疾患で回路の異常や過剰な興奮がどうして起こるのか、といったメカニズムの解明が進むことが期待されるとしている。









コメントをお書きください